| |
POLARIZATION VISION & TESTING IN ZEBRAFISH
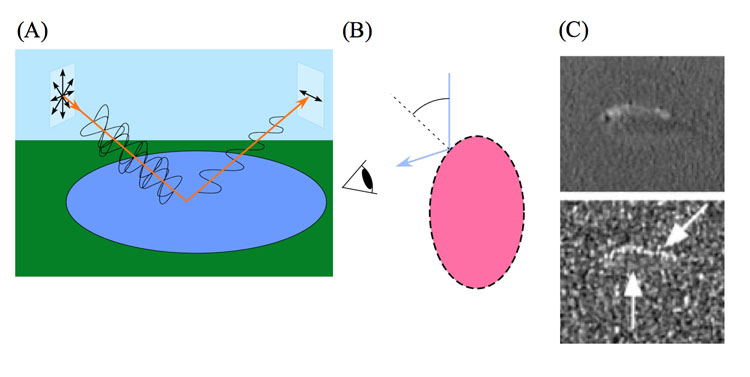
Probably due to our own perceptive limitations, the ability to sense ultraviolet and polarized light by other animals has long been underestimated, with respect to both its distribution and biological role. Polarization vision (PV) is the ability to distinguish between directions of oscillation (e-vectors) of a light beam. Human beings have a poor capacity to perform this ability (Haidinger’s brush; Fairbairn, 2001), in arthropods and many vertebrates like fish and birds it is a common feature (Horváth & Varjú, 2003) where it is often correlated with UV-sensitivity (Hawryshyn, 2010; Seliger, et al., 1994). PV may help the animal in orientation and navigation tasks (Weir & Dickinson, 2012) or to spot otherwise obscure objects (Cronin et al., 2003). The ability to discriminate between orthogonal e-vectors has been tested in different ways, varying from behavioural tests in intact bees (Kraft et al., 2011) and locusts (Mappes & Hornberg, 2004) to electroretinogram recordings in salmon (Hawryshyn et al., 2010) and butterflies (Stalleicken et al., 2006).
Inter- and intracellular structures like pigments that are located in visable regions of an animal (e.g. in the skin or behind transparent tissue) may selectively absorb or reflect e-vectors of the incoming light. This mechanism is used by cephalopods to produce polarized reflective patterns that could serve as an intraspecific communication-system among the colour blind but polarization sensitive cephalopods (Mäthger et al. 2009). The combination of polarized and non-polarized light that is reflected by respectively the black and white stripes of zebras, seems to disturb polarization sensitive horseflies, and as a consequence the zebras are less attacked by these biting insects (Egri et al., 2012). It should not be excluded that many more patterns with polarization modulation exist in nature. These may even have some adaptive significance, as they can only be perceived by polarization sensitive animals.
PV could play a prominent role in target detection by fish (Marshall & Cronin, 2011; Pignatelli et al., 2011). The underwater field of partially linearly polarized light hardly depends on the wavelength of the penetrating light (Cronin & Shashar, 2001). Especially in deeper water where colour vision becomes less reliable, polarization sensitivity may be incorporated in contrast enhancement, camouflage breaking, object recognition and signal detection and discrimination (Cronin et al., 2003). Although transparent zooplankton and insect larvae are strikingly birefringent (i.e. they can partially polarize light), their increased visibility to a predator with polarization vision primarily depends on other factors, like the polarisation of the horizontal background radiance and the partially depolarized light scattering from the prey (Johnson et al., 2011).
The ability of aquatic animals to discriminate between alternating e-vectors has been studied with different techniques, varying from conditioning of immobilized goldfish (Hawryshyn & McFarland, 1987) and free swimming damselfish (Mussi et al., 2005) to electrophysiological measurements in retinal tissue and optic nerve (Ramsden et al., 2008; Roberts & Needham, 2007). Using optokinetic responses, PV was studied in varies species of cuttlefish (Talbot & Marshall, 2010; Darmaillacq & Shashar, 2008), crayfish (Glantz & Schroeter, 2006) and salmonids (Hasegawa, 2007).
The optokinetic responses are elicited by the visual perception of a horizontally moving pattern of contrasting vertical lines (Jellali et al., 2005), whereby the eyes, head and entire body follow the moving pattern or whereby the eyes show optokinetic nystagmus (repeatedly following the movement direction of the pattern at about the same speed and then snap back in the opposite direction; Fritsches and Marshall, 2002). In many small animals the response is expressed by moving along of the entire body and is called optomotor response (OMR; see elsewhere on this site). The performance of optomotor response has been used to measure a number of thresholds, like motion detection (Kim & Jung, 2010), contrast sensitivity (Umino et al., 2008) and colour vision with respect to sexual selection (Rick et al., 2011) or UV wavelengths (Nava et al., 2011).
The advantage of the OMR behavioural paradigm is, that the same stimulus can be presented to an unlimited number of different intact animal species in order to observe possible PV. The underlying mechanism, however, is much harder to study with this technique. The relative simplicity of the set-up and the absence of physiological intervention, anyway, deserve to be considered next to the precise question and available equipment.
Apparatus & procedure
To test the presence of polarization vision, the OMR-apparatus was largely the same as described by Van der Meer (1994; see elsewhere on this site) and was also used to measure colour sensitivity differences in Pundamilia sp. (Maan et al., 2006). The central compartment contained the test object and consisted of a transparent cylinder of polymethylmethacrylate (transmitting most UV) with a transparent floor and a translucent white plastic cover. Below the floor a mirror was mounted under an angle of 45o to facilitate comfortable observations and/or recordings of the silhouetted test object. At the base outside the cylinder a movable ring was mounted to rotate in the plane of the floor and to carry replaceable cylindrical sheets representing the optical stimuli. The constant rotational speed of the ring (0,18 Hz) was powered by a 12V electromotor connected to the ring with an elastic rubber string. The following modifications were installed: one light bulb (Philips 60W, 220V) was mounted above the translucent cover and four lamps (2x Sylvania 11W and 2x Exo Terra Repti Gloo 2.0 13W; 6700K) with a spectral distribution approaching that of daylight were mounted at some distance around the cylinder to radiate through the surrounding demi-transparent sheets carrying the optical stimuli. The replaceable cylindrical sheets were:
- Control 1: a photographically produced vertical pattern of black and white lines of equal width (3,5 cm), to check the normal innate optomotor response.
- Test-carousel: connected slips (approx. 3,5 cm) of polaroid filters with alternating vertical and horizontal e-vectors, to test for linear polarization sensitivity.
- Control 2: slips (approx. 3,5 cm) of neutral density filters attached to one another and pressed between blanks.
The polaroid- and density filters were cut out of a PF006 (AFlash) polyvinyl alcohol-iodine sheet and a .9 GLLE211 (LEE) neutral density sheet respectively. The slips being 3,5 cm wide together with a rotational speed of 0,18 Hz produced a flicker frequency of 2,16 c/s which was amply below the measured flicker fusion frequency in cichlids (van der Meer, 1994).
Animals were tested according to the following procedure: when placed in the cylindrical tank of the apparatus, they were first exposed to Control 1 to check the ability to perform OMR. Unnoticed injuries of the visual system or brain damage may have deleted such an ability. It may also occur that the OMR is disturbed by unclear environmental annoyance (personal observations) or otherwise induced variability (Faisal et al., 2008). Therefore, measurements should be collected only after the animal is sufficiently adapted to its environment.
To ascertain the usefulness and applicability of the apparatus, polarization sensitivity was tested in common goldfish (Carassius auratus), a species of which it is quite certain that it possesses polarization sensitivity (Roberts & Needham, 2007; Hawryshyn & McFarland, 1987; Kleerekoper et al., 1973). Six specimens (body-lengths of ca. 5 cm) were obtained from a local pet shop and kept in a SuperFish Aqua 20 tank with an Aqua-Flow dual action filter (Aquadistri BV, The Netherlands) at room temperature (18-20oC).
Recordings were made during daily sessions of approximately an average of 60 minutes per specimen by measuring the absolute and relative duration of locomotion and by observing the direction of the locomotion in relation to the rotating stimulus. Accordingly, pecking at the wall, moving backwards, marking time by merely spinning around and gliding through, were considered as a standstill in the same way as resting behaviour. The directions of the stimulus rotation were alternately clockwise and counter-clockwise. The clockwise and counter-clockwise swimming movements were measured separately.
Only after a positive OMR had been recorded, the object was exposed to the Test-carousel and its movements were registered in a similar way as in Control 1. Afterwards, the collected data could provide information on the animal’s ability to perceive any modulation in the stimulus.
To complete the experiment, the object was exposed to Control 2. The purpose of Control 2 was to carry out a blank experiment with all parameters similar as in the test with the exception of the polarisation filters. Comparing the data of the test and Control 2 provided the significant evidence for the suitability and usefulness of the apparatus.
To generalize quantification of the responses to the rotation of black and white stripes, alternating e-vectors and neutral density filters, the number of seconds were measured that the object was moving in the same or in opposite direction as the rotating stimuli, during at least 600 seconds a time (with alternating clockwise and counter-clockwise rotations).
Two-tailed p-values were calculated with the Fisher’s exact test. The provided significant levels were used for the comparison of swimming movements in the same and in opposite direction of a stimulus rotation and to judge the optomotor response evoked by polarisation filters (Test-carousel) with Control 2 as the null hypothesis.
Testing the apparatus with goldfish
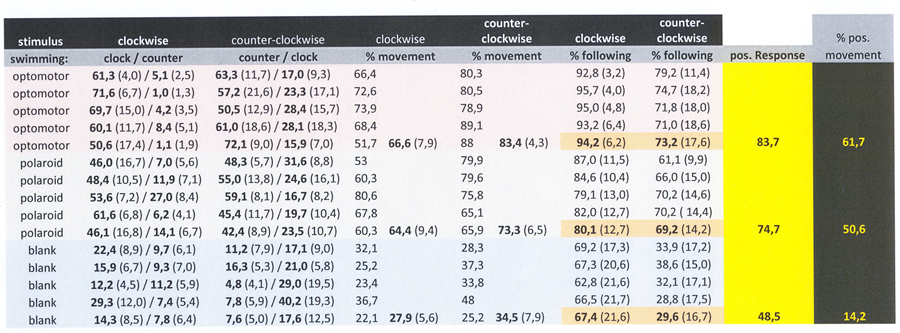
Exposure to the moving black and white line pattern evoked a vivid response in goldfish, dominated by swimming along with the rotating stripes (OMR), alternated with aggressive bites at the cylinder wall and swimming in opposite direction. Frequently, the test objects had a natural preference to swim clockwise(without stimulus), which was neutralized by the alternating clockwise and counter-clockwise rotation of the stimuli. The resulting OMR amounted 82,5% of the swimming movements with a standard deviation of 6,1 over 10 session of 600 seconds.
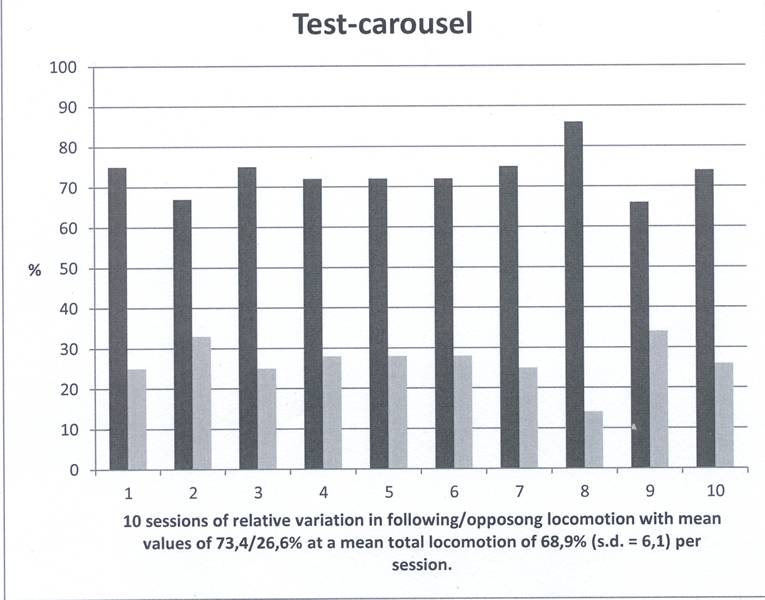
The Test-carousel with alternating e-vectors provoked slightly less swimming motion with a mean OMR of 73,4% with a standard deviation of 5,2 over ten sessions. The result of the test carousel was not significantly different from Control 1 (p=0.0597).
When exposed to the rotating blank, the fish were much quieter with a 45% swimming movement in the same direction of the rotation, again obtained from 10 measurements with a duration of 600 seconds each. Only in one out of ten sessions there was no significant difference between the Test-carousel and Control 2 (p=0,4615). The largest deviation from a 50/50 result during one session (being 38/62) was still not statistically significant (p=0.1169). The mean difference in swimming movements with the test carousel compared to Control 2 was highly significant (p<0.0001).
The highly significant following movements with the rotation of alternating e-vectors by goldfish indicated the usefulness and applicability of the apparatus to determine linear polarization sensitivity in small fish. The difference in OMR between Control 1 and the Test-carousel was traced to a different contrast in the presented stimuli. However, it should not be excluded that it was caused by a difference in effective quantum capture as movement detection is dominated by the larger long-wave sensitive cones (Schaerer & Neumeyer, 1996), and polarization vision is assumed to be primarily initiated by features of the smaller mid-wave sensitive cones (Roberts & Needham, 2007). Moreover, to evoke OMR the optimal spatial and temporal frequencies based on contrast need not to be similar as those frequencies based on polarisation (Maarten Kamermans, pers. communication).
The apparatus had also been used to demonstrate OMR in terrestrial insects (Acrididae) which are known to be polarization sensitive (Mappes & Hornberg, 2004). The locusts did indeed respond to the rotating alternating e-vectors. However, the movements were so different from the relatively regular swimming movements by fish that their response was not quantified and left out of studies in fish.
Testing zebrafish
Introduction
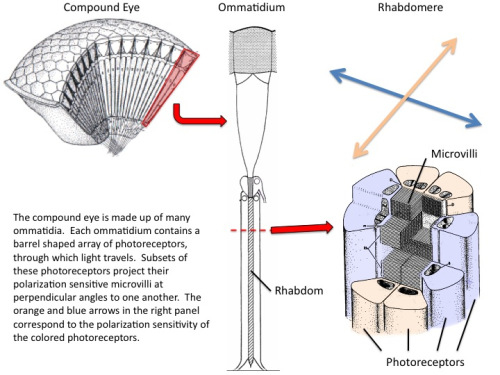
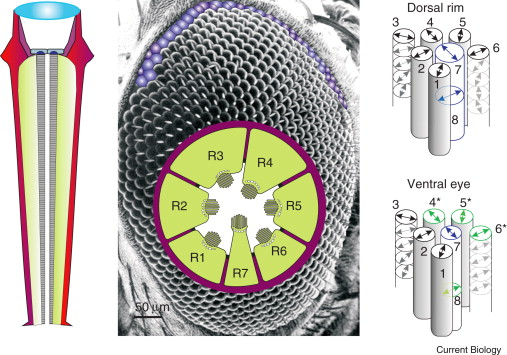
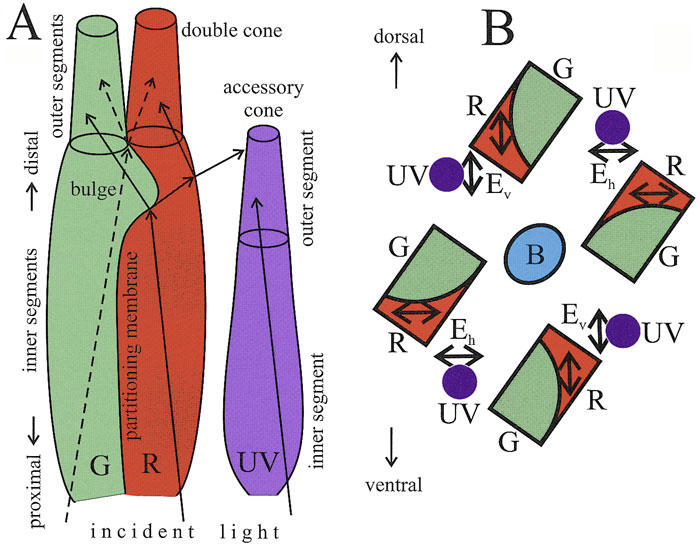
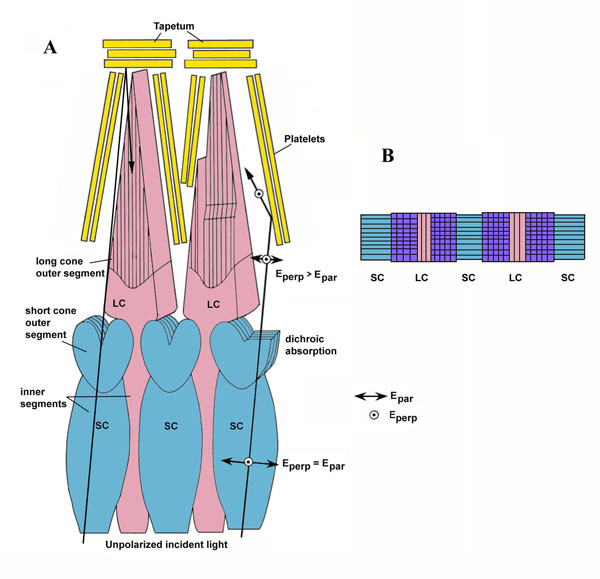
Zebrafish (Danio rerio), a cyprinoid teleost fish, has become an important laboratory model organism for many areas of research (e.g. Dahm & Geisler, 2006). For example, its visual behaviour and retinal structure have been studied in relation to ontogeny, genetics and human eye disease (e.g. Gestri et al., 2012; Gregory-Evans, 2012; Simmich, et al., 2012). The retina of the zebrafish includes four spectral types of cone photoreceptors, namely short ultraviolet (UV) sensitive and long blue sensitive single cones and fused green- and red- sensitive double cone pairs, which are aligned in parallel rows (Raymond & Barthel, 2004). In spite of detailed descriptions of the zebrafish retinal structures and visual behaviour, there is nonetheless “no evidence that zebrafish is capable to determine the e-vector direction of polarized light (Neuhauss, 2010)”.
By analogy to colour vision, PV can be a tool to distinguish different features of light. In the case of PV that would be the ability of a species to distinguish, at least to some degree, the planes of vibration of the electric vectors (e-vectors) of depolarized light (Cronin et al., 2003).
Results
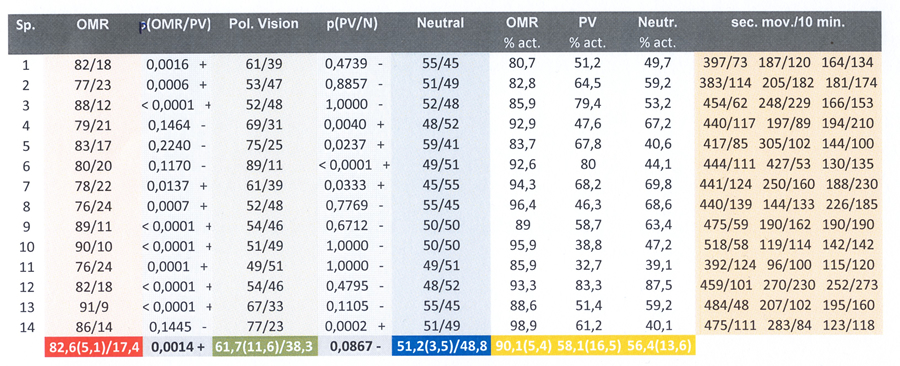
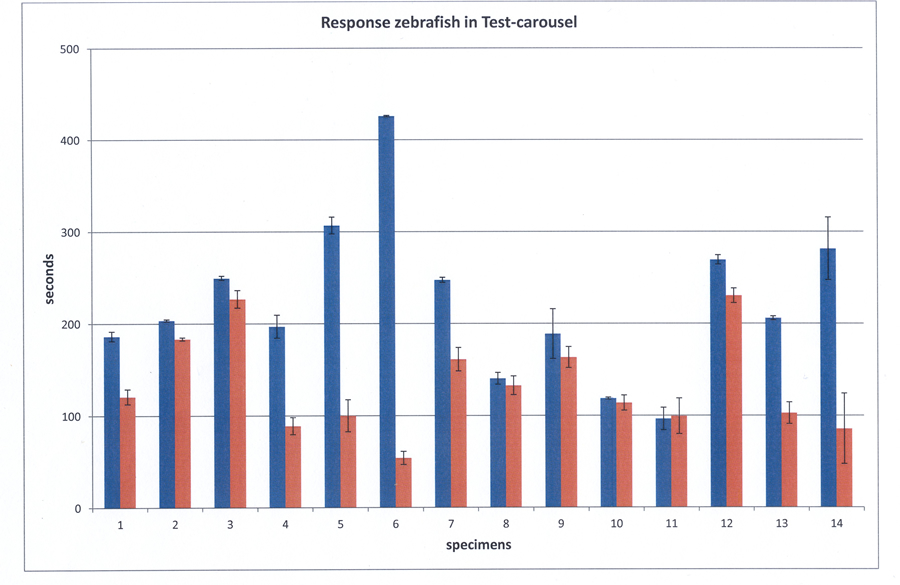
The zebrafish were constantly moving, even when isolated. Exposure to Control 1 did not immediately evoke a clearly recognizable OMR in all specimens. However, after a period of habituation (15 min) 14 out of 16 specimens swam significantly more often in the same direction as the stimulus, which was interpreted as a positive OMR. Two relatively small males did not perform an accurate OMR and were excluded from further analysis. The mean OMR amounted 82,6% of the swimming movements with a standard deviation of 5,1 measured over fourteen specimens.
The fish did not always swim at a uniform rate and the specimens performed some individual variations that were considered to be irrelevant for the purpose of this study, i.e. whether the species was capable to discriminate between alternating e-vectors. In spite of discrepancies in a number of specimens, statistical analysis did not reveal a significant difference between the mean response to the Test-carousel and Control 2 (p=0,0867) whereas a significant difference existed between Control 1 (OMR) and the Test-carousel (p=0,0012). The mean fraction following locomotion in the Test-carousel (with respect to all locomotion) amounted 61,7% (s.d.=11,6) and the mean positive response to Control 2 was 51,2% (sd=3,5).
Discussion
OMR could be instantly viewed in a qualitative way. However, for the purpose of a generalized quantification, more intensive measurements were carried out. The swimming direction could be generated by the rotating direction of the stimuli, without being a question of OMR, since the healthy fish were accurately equipped to perceive movement. However, when the swimming direction was evoked by OMR it clearly dominated the fish swimming behaviour.
The duration of the measuring sessions (10 min) was a bit arbitrary and came about by thorough observations. Short periods (1 min) gave too much fluctuations (very high standard deviations) and longer periods (30 min) provided little improvement, except for the object to become exhausted (pardon my anthropocentric interpretation).
It was a bit striking that in the Test-carousel practically all positive responses occured more often than the negative responses. If the swimming behaviour of the fish would have been influenced by any other visual stimulus than the alternating e-vectors or by the acoustic vibration caused by the motor driven rotating ring, this would have also been the case during the blank experiences. Comparison between the behaviour in the Test-carousel and Control 2 therefor eliminates such considerations.
A difference in perception between the human observer and the polarization sensitive fish should be considered as well. To a 'polarization blind ' observer, the rotating polarization filters will be perceived as alternating dark and light patches, only when looking through another polarization filter. Polarization sensitive fish, however, use different cones to detect differences in e-vectors. Their following response might therefore be a consequence of motion detection rather than optomotor response. This may involve different neuronal pathways.
Based on the mean outcome of the individual performances it is concluded that zebrafish as a species should be considered to be ‘polarization blind’. However, it should not be excluded that the discrepancies in some specimens are the result of true PV in these specimens.
There were no reports on structural differences in the retinas of adult zebrafish, although the cone photoreceptor mosaic appeared to change during ontogeny with respect to the cone subtype ratios and the precise row arrangement (Allison et al., 2010). It is possible that a more or less parallel positioning of double cones in the precise row arrangement provides enough intraspecific variation to discriminate between alternating e-vectors or not, depending on the average angle between neighbouring double cones in a row. Together with neighbouring UV-sensitive cones, this might provide a mechanism for PV (Novales Flamarique et al., 1998).
A completely different explanation for the occasional possibility of PV in adult zebrafish might be inherent to the protein-protein interaction within photoreceptor membranes (Roberts et al., 2011). Zebrafish are a simple object to point mutations (Streisinger et al., 1981) and together with the intense commercial breeding programs, the occurrence of ostensible insignificant proteins in some specimens of the wildtype line could have caused their PV. Due to a lack of ecological advantage this feature would not have evolved in their natural habitat. The possible capacity to discriminate between alternating e-vectors was performed by both males and females and could never be related to sex chromosomes as the latter are absent in zebrafish (Hofsten & Olfsson, 2005).
It was sometimes difficult to distinguish between general activity and swimming movements of the fish. Besides, the latter was often at a different speed than the rotating stimulus.To facilitate the observations and to reduce the degree of interpretation-freedom it is suggested to draw a polar-wedge diagram on the transparent tank-floor and count the number of segments that the fish crosses.< click here to go back to PAN-OPTICS & EVOLUTION |
|
|